18 On cultural learning and evolution: the selective advantage of altercentric participation?
Introduction
A form of cultural learning entertained without access to symbolic descriptions, conceptual representations and declarative memory has been identified to come into play by the pre-verbal mind. This is infant learning by virtual participation in the model's act and which in face-to-face situations entails sensori-motor mirror reversal (Bråten 1994). Such learning by altercentric mirroring comes into play at 9 months or earlier in the steps in children's development of intersubjective communication and understanding, distinguished in Bråten (ed. 1998):

These domains of intersubjective communion and understanding (specified in the right column of Table 18.1 below), remain operative throughout life. Hence they may be considered in the metaphor of a "staircase" with each lower-order step underlying and supporting higher-order steps (in contradistinction to the developmental "ladder" implied by traditional theories that assume egocentricity and lack of sociality from the outset), affording domains for various kinds of cultural learning, such as distinguished by Tomasello et al. (1993) and described in the following. In spite of pitfalls of drawing parallels between ontogeny and phylogeny, I shall also invite a comparison between these ontogenetic domains and the transitions in the evolution of culture from primate patterns to Mythos and Logos distinguished by Donald (1991) in his work on the 'Origins of the Modern Mind'. Representation and memory are key terms in his theory of evolutionary stages (listed in the left column of Table 18.1). The first stage involves a
transition from the episodic memory of social event perception in the primate-like culture to the invention of representations in mimetic skills for reproducing conscious, self-initiated, representational acts that are intentional but not linguistic. The second qualitative transition in Donald's account is from mimetic to mythic and oral culture involving semantic memory, and the third to a theoretic culture involving external symbolic storage and meta-linguistic skills.
I shall venture some speculations about the selective advantages afforded by learning by altercentric participation for the survival of hominid or human species in the phylogenetic period when ancestral infants could no longer ride on their mothers' back for protection and learning and before the invention of carrying bags of animal skin or other material. In that critical transition period, perhaps in the beginning of the Homo erectus period and before their inventing baby bags, infants and toddlers with an enhanced capacity for learning and listening by other-centred mirror-connectivity with distant facing adults would have had a selective advantage. The discovery of "mirror neurons" (Fadiga et al., 1995; Rizzolatti & Arbib 1998) indicates that in the neurobiology of the primate brain there appears to be a ground for systems that could have lend themselves to adaptations for mirror systems in humans that could support such infant learning by altercentric participation.
On Tomasello, Kruger and Ratner's theory of cultural learning
In approaches to cognitive development and enculturation there has been a gap between those who assume with Piaget the autonomy of the individual mind, and those who assume with Mead and Vygotsky the priority of interpersonal mediation. Tomasello et al. (1993) are to be congratulated for bridging part of that gap. Adherents of Vygotsky for the most part have been neglecting what the individual organism brings to the process of enculturation.
This Tomasello and his co-authors set out to remedy by focussing on the individual social-cognitive capacity for acquiring the skills and conventions of those around them. Cultural learning, they suggest, manifests itself in these three forms during human ontogeny: imitative learning (from 9 months), instructed learning (from 4 years), and collaborative learning (from 6 years). The authors consider these three types to reflect in the learner different concepts of person, as distinguished by Hobson (1987). As the child learns to understand others as intentional, mental, and reflective agents -- in that order -- new processes of social learning emerge. Imitative learning requires the concept of a person as an agent with intentions (zero order). Instructed learning requires the instructor to be understood as a mental agent with beliefs (first order), while in collaborative learning the other is understood as a reflective agent (second order) with the meta-capacity to reflect on own and others' intentional and mental states. In their theory, the underlying social-cognitive representations progress in these steps: from the processing of simple representation of the other's perspective in imitative learning, through self-regulative dialoguing or co-ordination of mental self- and other-perspectives in instructed learning, to, finally, integrated perspective-taking and intersubjectivity in collaborative dyadic learning.
Cultural nurture and learning embedded in communicative development
Tomasello, Kruger & Ratner (1993) compare ordinary children and children with autism with chimpanzees, and find grounds for considering cultural learning a unique human form of social learning. The authors do not specify, however, how cultural learning depends on prior primitive capacities in the learner, even though partly implied by the rich comparative evidence offered. They have left unexamined, however, the socio-emotional and auto-operational roots which the infant brings to bear on these processes.
The capacity to enter into an engagement of feeling with others in a direct, nonreflective sense with bearings for cultural learning is manifested in early infancy. Normal human infants engage actively and effortlessly in mutually attuned affective contact with adults (Stern 1985; Trevarthen 1990a). Even in peer contact, when opportunity arises, 3-month-olds have been found to engage in reciprocal gazing, smiling, vocalization, and sometimes reaching for each other (Field 1990). Such capacities, indicative of sociality from the outset, may be allocated to the first of several steps in early communicative development, and which continues to underlie and support higher-order abilities later in life.
I shall point to a form of cultural learning which may be operative in the preverbal infant soon after birth and, hence, differ from advanced learning by the verbal mind with access also to symbolic descriptions, conceptual representations and declarative memory.
A pertinent example of cultural learning right from the beginning has been documented in studies of speech perception by preverbal infants. Kuhl (1998) reports how infants during the first 6 months of life learn to tune their speech sound perception to the music and sounds that make sense in the native language of the culture into which they are born, discarding sounds that make no sense in that language. Another example of early cultural learning is my own recordings of how 11-month-olds can reciprocate their caregivers' spoonfeeding, requiring what I term altercentric capacity to learn by virtually moving with the other's goal-oriented movements (Bråten 1996c, 1997a, 1998a).
Such interpersonal communion and learning by altercentric participation underlie and nurture the emergence of first-order interpersonal communication mediated by symbols and conceptual representations. And furthermore, without the natural capacities and cultural nurture afforded in the primary domain of interpersonal communion, second order processes of meta-understanding of others' understanding can hardly be realized.
Second order mechanisms, whether studied from the viewpoints of mindreading, theory of mind, or simulation of processes in and between minds, may be evoked in this domain upon breakdown of intersubjective understanding during symbolic conversation. Some researchers (e.g. Astington et al. 1988; Whiten 1991) attributes to children around 4 years old to have formulated a theory of others' minds enabling understanding of others' mental states, while others (like Bråten 1974, 1998; Gordon 1986; Harris 1991) rather consider children or adults to use their imagination to mentally simulate others' thoughts and emotions.
While finding "theory-of-mind" findings useful, Tomasello et al. (1993:502) prefer the simulation-of-mind account. They point out that what they have described as perspective-taking could easily be described in terms of simulating the intending and thinking of another person, provided that this does not rest on the assumption that children have to understand their own mental states in order to simulate those of others.
This is consistent with my own view of co-actor (coding) simulation processes evoked in conversation (cf. essays 4 and 5, this volume). However, as has been indicated, there are socio-emotional and bodily alter-centred processes that are not just precursory, but continue to underlie and play a role also in such higher order circular self-other processes. Altercentric participation in the other's movements has been described in terms of a virtual other mechanism which complements the infant's bodily self feeling. This operational notion, I submit, affords an explanatory account of the bridge between intra-personal mechanisms and inter-personal processes in the various COM1M2 domains listed in table 17.1. The virtual other mechanism allows for altercentric perception and completion of the interlocutor's intention in first-order domain (M1) of symbolic conversation, and for mental simulation of the other's mental processes in the second-order domain (M2) when intersubjective understanding breaks down. This, then, provides a clue to advanced modes of perspective-taking, asserted by Tomasello et al. to be required in advanced forms of cultural learning.
A comparative perspective: communicative development and cultural evolution
Advanced cultural learning is not just characterized by perspective-taking in virtue of symbolic representations, but by recursive mental symbolic simulations of self- and other-perspectives in dialogue (Bråten 1973a, 1982b; 1988). The developing mind emerges as dialogical, perhaps the unique operating characteristic of human consciousness. Asymmetric (instructed) learning in which the instructions are re-enacted in overt self-talk, and the evocation of internal dialogue as self-guiding, not just re-enacting mentally the other's instruction, but co-creatively engaging in symmetric (collaborative) learning (Tomasello et al. 1993), which clearly occurs in peer interaction at four years. This is the age in which perspectives may be simulated by virtue of models of others' minds, even when the child attributes to the other a belief which the child knows to be false. That presupposes the capacity to hold concurrently not just two different perspectives in mind, but somehow to simulated the other's mental state of belief.
Even pretence, arising earlier, requires the capacity to entertain in parallel two different perspectives and being aware of which of them applies to the actually present other. The ability to engage in internal dialogue between one's own perspective and another attributed to an absent other appears to be unique to humans and depend on a higher form of sociability, albeit necessarily nurtured by experiences from interacting with actual others in prior presentational mediacy.
Comparison with semi-enculturated chimpanzees
Tomasello et al. (1993) attribute to humans species-specific social-cognitive abilities to engage in cultural learning, entailing intersubjective understanding of others as mental agents. They find no evidence of this ability in our closest primate relative, the chimpanzees, even when raised in human like cultural environments.
In their comparison of children and chimpanzees, Tomasello et al. (1993:508) find that there is something acultural about the learning and social lives of chimpanzees. In spite of this, enculturated chimpanzees are found to manifest imitative learning and to demonstrate simple perspective-taking skills. Higher types of cultural learning and the ability to teach depend according to the authors on the understanding of others as mental agents. The chimpanzee, even when raised in human like culture, is in their view without this capacity. Chimpanzees, they claim, can neither teach nor internalize the instructions of others. This is contested by Kim Bard (1993:511-12) who refers to examples to the contrary (see also Bard in Bråten (ed.) 1998).
Tomasello et al. (1993: 509) point out that advanced self-regulated cultural learning, beyond the capacities of children with autism and enculturated chimpanzees, involves recursive intersubjectivity and internal dialoguing between mental perspectives, including a virtual other perspective. True, provided prerequisite cultural nurture, the learner's virtual other, in virtue of complementing the bodily self and having the operational efficiency of an actual other, serves inter alia later in ontogeny the inner observing function described by Tomasello. But equally relevant, posited as inherent, not constructed, albeit changed with cultural nurture, the learner's virtual other mechanism permits the learning mind to recreate and transform itself in the form of the dialogical, whether in (i) in self-engagement (with the virtual other), or in (ii) affectively tuned dialogic engagements with actual others, including peers. In this way this inner companion perspective, complementary to the bodily self, allows for learning by altercentric participation in models' movements affording e-motional memory for bodily circular re-enactment. This is not just a common base for the cultural learning mechanisms examined by Tomasello and his co-authors. Serving as an companion perspective in inner dialogue and self-conversations, the learner's virtual other mechanism provides a basis for the ontogenetic leap to higher-order virtual pre-enactment of others' acts and mental processes later in ontogeny, i.e. to the nurtured rise from bodily re-enactments in felt immediacy to mental simulations of other minds in representation mediacy.
Communicative development and (cultural) evolution compared
Tomasello et al. (1993:509) regard the primary evolutionary context in which primate cognition evolved to be a social one and, hence, that most of its unique features stem from adaptations to social challenges such as communication, social learning, cooperation, as well as competition and deception, in which primates are unique social strategists:
"Our contention is simply that in this context human beings evolved species-specific social-cognitive abilities to understand the psychological states of conspecifics in terms of their perceptions and intentions, their thoughts and beliefs, and their reflective thoughts and beliefs, which allowed them to take the perspective of others and to participate with them intersubjectively." (Tomasello et al. 1993:509)
Such processes of social cognition, they claim, led humans to the species-specific ways of learning from one another in cultural learning, which then kicked off the evolutionary and historical processes that led to human cultures.
On the origins of the modern mind
In his 'Origins of the Modern Mind', Donald (1991) distinguishes in phylogeny the cultural stages of episodic immediacy, resembling patterns found in the great apes, and Mimesis from the succeeding symbolic stages of Mythos and Logos. With partly a focus on different kinds of memory involved in the evolving cognitive and cultural patterns, Donald describes transition stages in terms of these distinctions (cf. the adapted expressions in Table 18.1, left column):
I. Episodic (primate) culture, in which social event perception and self-reproduction of communality rely on episodic memory of concrete events
II. Mimetic (homo erectus) culture, reproducing itself by individual and collective mimesis, i.e. imitative and creative re-enactments of standardized gestures and tool-inventions involving procedural memory
III Mythic (archaic human) oral culture, involving pictorial carvings and speech, relying on semantic memory
IV Theoretic (modern human) linguistic culture, evolving external symbolic storage systems and meta-linguistic skills.
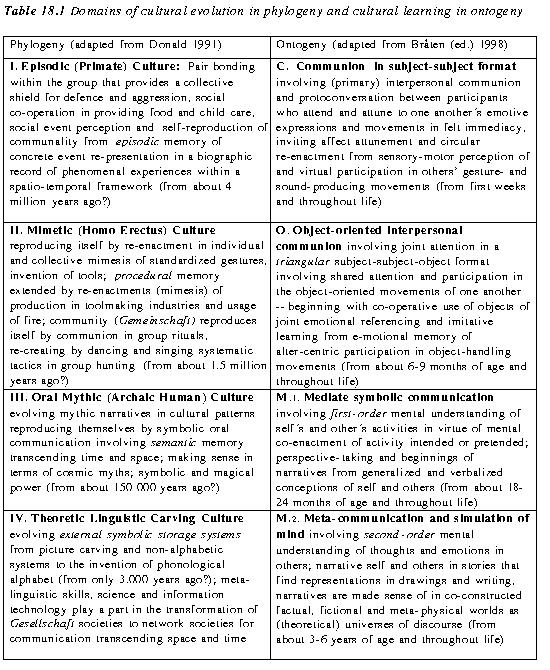
They invite comparison with the ontogenetic domains of children's communicative development distinguished above (Table 18.1). However, even though auto organizational inventiveness plays a part in phylogeny, sociogenesis, and ontogeny, they differ in this critical respect: While societal evolution from phylogeny has involved the generation of new cultural lifeworlds in new domains never before entertained, the child is born into the peculiar rhythms and musical sounds of a language culture that already exists, nurturing the self creative transitions from consensual communion to symbolic, narrative communication. While the child is learning to "swim', as it were, in the existing cultural 'ocean' into which the child is born, the phylogenetic transitions described by Donald entail the creation of cultural 'oceans' that never before have existed. Furthermore, the meta-communicative simulational capacity (M.2), entailing virtual participation in conversational partners, however, must have evolved with the advent of the mythic culture (III), if not earlier.
In spite of such pitfalls and lack of correspondence, there are also promises in drawing parallels between phylogeny and ontogeny, as demonstrated a century ago by Baldwin (1895). One possible parallel is this: Advanced levels in both ontogeny and phylogeny are probably supported by processes at primary and primordial levels, continuing to be operative and to underlie higher-order abilities and forms, even though disguised from and ignored by the egocentric and ethnocentric observer.
Donald emphasizes the crucial role of systems for representation in cultural evolution:
"Human did not simply evolve a larger brain, and expanded memory, a lexicon, or a special speech apparatus; we evolved new systems for representing reality. During this process, our representational apparatus somehow perceived the utility of symbols and invented them from whole cloth; no symbolic environment preceded them." (Donald 1991:3).
True, but environments marked by traces of others and of other things, recognized by proto-declarative pointing as signs of others (footprints of hunters or drops of prey) who had been there, would have preceded the invention of such symbolic re-presentations. And the imitation and imitative learning from procedural memory entertained in the mimetic (homo erectus) culture would have contained signature elements of re-presenting the model or model act enacted in the past.
This pertains to the transition to a mythic (archaic human) oral culture, involving pictorial carvings and speech, and coming to rely on semantic memory. Now one may ask about the kind of leap entailed by the transition from the episodic (primate) culture, relying on episodic memory of concrete events, to, involving imitation and imitative learning from procedural memory.
Adaptive problems of childcare and early learning in early human evolution?
The cogent question is this: What kind of selective advantages may have been afforded by the emergence of mechanisms of mimesis, as described by Donald? With reference to the kind of infant capacity enabling pruning in infant speech perception before 6 months (Kuhl 1998), deferred imitation by 9 months (Meltzoff & Moore 1998), and learning by altercentric participation in face-to-face situations by 12 months (Bråten 1998), I offer this conjecture:
Infants and toddlers with an enhanced capacity to learn to cope by watching and listening to adults would have had a distinct selective advantage, having to be grounded, in the critical transition period before the supposed invention by Home erectus of baby carriers (by animal skin or other material).
On the origin of humankind
In the middle of 1980's Richard Leakey and his colleagues uncovered the entire skeleton of a boy in East Africa, who must have died at the edge of an ancient lake more than 1.5 million years ago. This Turkana boy, a member of the species Homo erectus, has inspired Leakey (1995:93-98) in his account, The Origin of Humankind. On the basis of the archeological evidence that he and others have uncovered, he attempts a re-construction of a scene that may have applied to some members of this species, gathered at a stream-side campsite:
"[w]e see a small human group, five adult females and a cluster of infants and youths. they are athletic in stature, and strong. They are chattering loudly, some of their exchanges obvious social repartees, some the discussion of today's plans. Earlier, before sunrise, four adult males of the group had departed on the quest for meat. The females' role is to gather plant foods, which everyone understands are the economic staple of their lives...
Three of the females are now ready to leave, naked apart from an animal skin thrown around the shoulders that serves the dual role of baby carriers and, later, food bag.." (Leakey (1995:93-97)
In this re-construction, they are attributed the invention of a combined baby carrier and food carrier, using skins of animals they have butchered with their tools. Compared to earlier hominid species they have larger brains, and probably a longer childhood entailing a long period of learning and growth, very unlike the progress in great apes which is almost directly from infancy to adulthood. Biological anthropology informs us that human infants, compared to other primates, are in a biological sense prematurely born as a consequence of our large brain demanding growth outside of the womb due to the limits of the human pelvis adapted to bipedal gait and running. This suggests that this prolonged period of childhood may have played a pivotal role in the biological and cultural evolution of humankind.
How would infants fare when they could not ride on mothers' back and before carrying bags?
Now venturing back a little further than the scene described above, before the invention of carrying bags by using animal skin or other material, how would the infants and toddlers fare, having to be let down on the ground? The cranium Walker and Leakey (1978) found at this same lake Turkana in Kenya in 1972, appears to represent the genus Homo (Homo habilis) perhaps two million years ago. They may have been gatherers and scavengers, perhaps not hunters, using stone tools, but probably not tools for baby and food carrying.
Living in the open savanna with few trees to climb for safety from predators, a hominid mother would have difficulties in climbing a tree while holding her infant, who unlike the offspring of apes probably was unable to cling to her by own force. She would be dependent on her kin-based band for the protection and shared caretaking of her children. Vegetation she could gather, but only with one hand, and she would have to let down her toddler on the ground as she sorted or worked with the food. Both females and males, when not out foraging, gathering plants or insects or looking for a carcass left by predators, would have to watch the children, protecting them from a variety of dangers, not just predators; they were vulnerable to a number of parasites and pathogens. Children were few and took a long time to learn to take care of themselves. Without efficient direct infant-adult communication and mediation of coping procedures from adults to children, survival would be difficult. In such critical conditions those would be favoured who entertained a leap to a mental architecture that increased the efficiency of direct face-to-face communication and of offspring capacity to learn by intergenerational transmission.
This is a defining characteristic of culture, albeit perhaps not unique for humans (cf. the appendix A1, this volume). Wild chimpanzees living today in West and Central Africa appear to exhibit significant variations, suggestive of culture.2 Chimpanzee offsprings are born strong enough to cling by own power to their mothers' fur, and when older the infants ride on their mothers' back. Carrying their young offsprings in this manner by virtue of their quadrupedal gait, great apes are able to keep their infants safe, while affording them the experience of orienting themselves in the world from the mother's physical position, moving with her movements. If this way of having the infant hold on to mother was the mode of ancestral apes in Miocene time, then it could no longer be upheld by hominids who had risen to their back limbs, entailing skeleton changes and other adaptions that could accommodate a bipedal gait for efficient locomotion in nonwooded savanna environment.
Killackey (1995:1244) point to these possible advantages of the shift from quadrupedal to bipedal gait: energy efficiency, raising the eyes for a better view over the savanna grasses, and freeing the forelimbs for other functions. Possibly true, but at least one of the mother's arms would have to be used to carry her infant, restricting her foraging, and when squatting to handle the food with both hands, she would have to put the child down -- aside or in front to keep an eye on the child, perhaps ready to vocalize in a way that could alert, alarm or sooth the infant as the need might be. As for the infant, deprived of the safe learning by moving with the mother's movements on her back, the child would now have to stay tuned to her vocalizations and to learn from watching her and the others from the ground.
While even neonates of great apes are able to physically cling to their parents for safety and in silence, the Homo neonate would have to resort to a sort of early "mental clinging" (Bowlby 1969), entailing an evolved mental architecture for interpersonal connectivity that becomes operative even in face-to-face relations at some distance. Taking after the facial expressions, vocalizations and gestures of the other is a way to ensure such mental connectivity in such settings, while being preparatory for learning by imitation. The emerging mechanisms for what Donald (1991) specifies as Mimesis and attributes to Homo Erectus (from about 1.5 million years ago, when the Turcana boy is dated) in early Pleistocene time, and the kind of interactions that Leakey (1995) describes in his scene re-construction, may possibly have evolved as intergenerational means of early child learning in the scavenger-gatherer and hunter-gatherer society.
The conditions for scavenger-gathering hominids, perhaps even for early Homo erectus hunter-gatherers before they may have begun to use animal skin or other material to carry infants, animals and plants (as suggested by Leakey (1995:93-98)) may have entailed constant threats to adults and offsprings by toxic plants, predators and other forms of hazards. In such hazardous condition species survival would depend inter alia on child care and children's capacity to learn by watching and listening to the adults. Unlike the great apes, with offsprings riding on the back of their parent and learning to move and operate in the world from that position, these children may have had to depend for their survival on a kind of alert and attentive face-to-face engagements with the adults, listening to them and learning from them even when not being hand-guided. (Figure 18.1).
[Figure 18.1 about here and with this text on the same page, or the opposite page:]
Figure 18.1 Could this be a scene typical of the early scavenger-gathering hominids, perhaps even for Homo erectus hunter-gatherers before they may have begun to use animal skin or other material to carry infants, animals and plants? If this were a typical scene, in which adults and offsprings were constantly threatened by toxic plants, predators and other forms of hazards, child care and 'baby sitting' would not suffice without children's capacity to learn to cope by watching the adults. Unlike the great apes, with offsprings riding on the back of their parent and learning to move and operate in the world from that (egocentric) position, these children may have had to depend for their survival on a kind of alert and attentive face-to-face engagements with the adults, learning from them even when not being hand-guided. If that were the case, then those with a capacity for other-centred mirroring and, hence, for what I term learning by altercentric participation would have had a distinct selective advantage.
If that were the case, one may speculate about the possibility that the advent of infant mechanisms for other-centred mirroring and, hence, for what I term learning by altercentric participation (Bråten 1997ab, 1998ab) would have been a selective advantage in such conditions, favouring generation after generation over early human evolution.
Mentioning these speculations to Fredrik Barth, he told me about a pattern he had observed in central New Guinea when studying the Baktaman (Mountain Ok) in the 1960s (Barth 1975):
"In this inner New Guinea community, the mother would remain in the birth hut/menstrual hut for approximately the first three months after giving birth, holding the infant in her arms. During this time, the baby was not wrapped in any form of cloth or covering, but held against the mother's skin -- the mother herself being uncovered apart from her short grass shirt. Only after the child was regarded as sufficiently robust would the mother start moving around, carrying the baby on her back in a string bag. At this point, the baby would also start sometimes being placed briefly on the ground, nested in the string bag. Though Baktaman women had, and generally carried, a tapa cloth head-and-back cape, this cloth material was not, as far as I observed, used to wrap the infant/baby." (Barth, personal communication 1999).
Indeed string bags, and perhaps also animal skins for carrying, may have been a late invention, perhaps in the later part of the Homo erectus period (cf. Leakey 1995:93-98). What Barth and I find almost inconceivable is how, before the advent of such inventions, Homo erectus offspring-care during the first two or three years after birth could be coped with -- even though there may have been elaborate social group structures for shared responsibilities and "baby sitting". Rather than placing all the emphasis on selective pressures on the adult caretakers, given the long period of infancy development outside the bipedal mother's womb, I venture to suggest that infants and young children with an enhanced capacity for early and rapid imitative learning by face-to-face observation would provide a distinct selective advantage in evolution.
One may even speculate whether it may have furthered also some kind of Baldwinian effect (Baldwin 1902) in the phylogeny of warning signal and sound perception.3
Cosmides and Toby (1995:1208) describe the ancestral hominids inter alia as a long-lived, low-fecundity savanna-living primate species with a sexual division of gathering and hunting, involving co-operation and making tools. They were mammals with altricial young, with an extended period of female investment in pregnancy and lactation, with pair-bonds and long periods of bi-parental investment in offspring. They were vulnerable to a wide variety of plant toxins and predators, parasites and pathogens. If such were the conditions of a hominid species that may have prevailed for one million years having relatively few offsprings with long immaturity and exposure to dangers that required a lot of adult caretaking, we should perhaps regard infant-adult and child-adult communication to pose an adaptive problem perhaps as important as for example food source locations. Gathering and hunting may have profited from an evolved mechanism for allocentric mapping and perception that complemented mere egocentric or body-related mapping. The kind of alert and attentive face-to-face engagements called for by adults and infants may even have entailed the evolvement of mechanisms for other-centred perception and, hence, for what I term learning by altercentric participation. Threatened by toxic plants, predators and other forms of hazards, in generation after generation over hominid evolutions and before the invention of bags for carrying infants and toddlers, the capacity for such other-centred mechanisms for mirror-connectivity would have contributed to survival.4
Hominid species may have been faced with extinction, I submit, if their young offsprings were unable to cope with face-to-face learning and listening by altercentric mirroring.
Exposure of infants, toddlers and young children to hazards (predators, parasites, poison, etc.), may have favoured those that had an enhanced capacity for rapid learning by observation and listening in face-to-face settings. Infant and early child adaptations for efficient learning by altercentric perception and participation in facing adults' activities may constitute the phylogenetic roots of the kind of reciprocal mirroring we may observe in early human ontogeny, and indeed throughout normal human life, for example in the intimate dialogue between two teenagers.
And in the primate neurobiology there appears to be a ground for systems that would have lend themselves to adaptations for systems in humans that could support such altercentric participation. This is the occurrence of so-called "mirror neurons" discovered in the primate brain.
The discovery of mirror neurons in the macaque monkey brain
Mirror neurons have been found in the macaque monkey brain to discharge both when another is observed grasping a piece of food and when the monkey is preparing for grasping the piece by itself (Di Pellegrino et al. 1992). On the basis of studies of human subjects, Rizzolatti and Arbib (1998) refer to evidence which suggests that in humans a similar mirror system may be allocated to Broca's area, which not only serve speech, but appears to come active during execution and imagery of hand movement and tasks involving hand-mental rotation.
The group of neurophysiologists behind this discovery, headed by Rizzolatti, has also uncovered another class of neurons, which they term "canonical" neurons. They become active at the sight of a graspable object, while mirror neurons become active upon observation of another individual grasping the object. Vittorio Gallese, a member of this research group, describes their discovery and some of the implications in this way:
" Both canonical and mirror neurons become active motorically when a given movement pattern of the hand is aimed to a certain target in a certain way to achieve a certain goal. These two types of premotor neurons respond also to visual stimuli in a peculiar way: canonical neurons respond to the observation of graspable objects, even in the absence of any movement of the monkey.
Mirror neurons do respond to the observation of hand actions, performed by other individuals, which are congruent in terms of goal with the motorically coded ones. Similar results have been obtained also in humans with neuroimaging techniques.
Within this context, a goal could be conceptually defined as the explanation in teleological terms of a "willed" relational attitude [...].
The same relational attitude is applied when observing other behaving individuals. The observer begins to "understand" the observed behavior of a third party when this process of "motor equivalence" between action observation/execution is established by means of a shared motor representation." (Gallese, personal e-mail communication 1999).
The mirror neurons discovered in the macaque monkey allow, I presume, for coding in terms of goal in a way that is congruent with the coding in terms of goal allowed by canonical neurons, i.e. in both cases, I take it, the monkey relates from the monkey's stance to the object and to the individual grasping the object.
When we turn to humans, however, I expect that there will be uncovered a difference in the way which the nervous system elaborates a code that "classifies" the objects of the external world according to their relational value for the acting subject, and the relational attitude applied when observing other behaving individuals -- at least when occurring in face-to-face situations. In the infant learner this would somehow presuppose altercentricity on the part of the learner to acquire a shared motor representation from the experience of feeling to virtually co-enact the execution observed in the other. In face-to-face exposure a perceptual mirror reversal is required for the "translation" into a proper premotor programme for circular re-enactment by the learner.
A forthcoming meeting on mirror neurons and the evolution of brain and language
Also, the human conversation partner who virtually participates in the speech listened to by mirroring what she hears and sees the dialogue partner to be doing, is able to do so, I have posited, not by merely mirroring the other from her own (egocentric) stance, but by a virtual circular reversal enabling her to simulated the speech processes from the other's stance. The key feature here is speech perception, studied in preverbal infants by Kuhl (1998) and which may indicate that children perceive speech by overtly or covertly co-enacting the vocal-oral movements that produced them rather by an auditory processing of the speech sounds. This may be similar to what occurs in an intimate verbal conversation in which the adult listener does not perform an acoustic analysis, but rather unwittingly mirrors the speaker's process of producing the utterance, post-dicting, as it were, the oral-vocal movements (cf. Bråten 1974; Liberman 1957, 1993 -- referred to also by Deacon 1998:359). This entails speech perception by altercentric participation in the other's speech production. Kuhl (1998) finds that already by six months infants have narrowed their speech perceptual space to become indifferent to sound distinctions that make no difference in the native language. This may perhaps involve the mechanism of learning by altercentric participation in familiar sound productions, long before acquisition of meanings.
Enculturated chimpanzees can learn some sign language, while lacking this capacity to move in the "other shoes", as it were, from the other's stance. I expect such participation by sensori-motor reversal to be unique to humans, perhaps evolving from selective pressure on infants to learn from face-to-face interaction (unlike offsprings of nun-human primates, riding on their back and facing the world from the same direction).
Examining the experimental results suggesting that a mirror system may be operative in humans, Rizzolatti and Arbib (1998) refer to Donald's (1991) assumption about mimesis as precursor to language, and speculate on the sequence of events that might have led from gestural communication to speech. It is likely, they state, that the human capacity to communicate beyond that of other primates depended on the progressive evolution of the mirror system in its globality. We shall have the opportunity to pursue such questions at an international meeting primo July 2000 on "Mirror Neurons and the Evolution of Brain and Language". In their conference announcement, the organizers Vittorio Gallese and Maxim Stamenov point out that the discovery of mirror neurons sheds new light on the neural correlate of social sharing of intentions and actions. The action of mirror neurons seems to provide a bridge and mapping between one's own behavioural actions (and their potential monitoring) with the observed actions of other individuals. In the development of the language faculty, they suggest, there may be both phylogenetic and ontogenetic links between behavioural action, gesture and communicative verbal action, and the discovery may provide a neurobiological basis to account for the emergence of language, based on these assumptions, listed by Gallese and Stamenov (1999):
(a) Language skills have emerged through evolution by means of a process of pre-adaptation in which specific behaviours and the nervous structures supporting them, originally serving other purposes, acquire new functions that side and eventually supersede the previous ones.
(b) With the primate premotor cortex as the common ground, an evolutionary continuity to language skill may be traced from pre-language arm- and hand-manual behaviours.
(c) The specialization for language of human Broca's region appears to emerge from an ancient mechanism, the mirror system, originally serving action understanding.
Some qualifications and added assumptions
Collaterals to b) and c) embedded in language terms 'comprehension' and 'begreifen'?
I find the above assumptions plausible, and see them partly reflected in the 'wisdom' or metaphors reflected by some contemporary languages, such as English, French, Russian and the Scandinavian languages. They have all have words indicating a link between the manual and the mental acts of grasping. For instance, in French the term 'comprendre' for comprehend has the root sense of grasping, 'prendre'. In English the term 'prehension' has the double meaning of 'act of grasping' and 'mental apprehension'. In German the term for conceptual understanding is 'begreifen' with the root term 'greifen' which again means grasping by hands. We find the same reflected in Danish, Norwegian and Swedish. In Russian the corresponding terms for the verbs comprehend (oxvativatj) and manual grasp (sxvativatj) have the common stem word 'xvatatj' which means grasp.
Precursory to current systems serving altercentric mirroring in phylogeny and ontogeny
The above assumptions (a - c) clearly pertain to the topic of the present essay (and to essays nos. 16, this volume). In view of my independent identification of altercentric participation and learning by altercentric mirroring in early ontogeny I would like to add these qualifications and supplements:
(d) While an evolutionary continuity to language skill may be traced from pre-language arm- and hand-manual behaviours, there appears to be such a continuity also in early ontogeny; i.e. from manual action learning and understanding to conceptual comprehension and mental understanding.
(e) While the phylogenetic specialization for language appears to emerge from an ancient mechanism, the mirror system, originally serving only early action learning and action understanding in human evolution, that system is precursory to systems that continue in homo sapiens sapiens to subserve mechanisms in preverbal learners and verbal dialogue partners for altercentric mirroring and participation.
The mirror system has been located to involve human brain areas that subserve speech and mental hand rotation. It has been recorded to be evoked in human subjects when another hand-grasping individual was observed and when the subjects imagined themselves grasping the object without actually moving their hand. I find it highly pertinent that Broca's area which may be involved in the mirror system also become active in tasks involving mental hand rotation.
It obviously pertains to the kind of altercentric mirroring which may be attributed to intimate dialogue partners who mirror one another's postures and gestures and sometimes complete one another's utterances. And it pertains to preverbal learning by altercentric participation which must have been a distinct selective advantage in phylogeny.
On further differences in non-human and human primates
A pertinent question is this: Would the mirror system found in human subjects differ in terms of orientation from mirror neurons found in nonhuman primates? They are probably similar in the sense of transcending egocentricity by the fact that observation of goal-directed acts by other individuals give rise to firing as if to do the same act.
But I expect the systems to differ in terms of other-orientation, with altercentricity probably unique to the human system. Unlike human infants and toddlers who during some critical period in human evolution had to cope by face-to-face learning and listening, the infant offsprings of no-human primates ride on the adults' back and experience the world and events and others in it from that egocentric and safe position. By turning allocentric the mirror system in grown-up non-human primates transcends that egocentricity, but I do not expect it to be altercentric like the mirror systems in humans which, I submit, would have had to become by necessity fundamentally other-oriented for the human offsprings to survive in the critical period before baby carriers were invented. There would have been a definite selective advantage to early humans, not to non-human primates, of a kind of mirror system which not only served to mirror the act of another individual, but which enabled virtual participation in that act as if jointly co-enacted and, hence, learning to cope by altercentric participation.
Thus, I expect mirror systems in human to have, when sensitized, the unique property of enabling mirroring the other's movements from the other's stance. This enables learning by virtual participation in the other's performance from the point of view of the other even when facing the other. The evolved capacity for this kind of participatory attunement to the gesture and sound producing movements and object-handling of models and teachers has favoured the infants and young children of our surviving ancestors. In this, I attribute much of the survival value to infant capacity, rather than to adult child care under such critical ancestral conditions. .
I assume that the evolved capacity for altercentric perception and participation would have been adaptive for the intergenerational transmissions in the mimetic culture of Homo erectus. If that is tenable, then the posited virtual other mechanism, complementary to the bodily self, which enables such mirror connectivity to others, may be reflected already by the Homo erectus brain architecture. We should expect the ancestral human brain to have an asymmetry that enables the realization of a companion perspective complementary to the egocentric body (schema) perspective, perhaps in the chiral (mirror image) manner that right and left hand are related.
According to Killackey (1995:1251) the functional asymmetry of the human neocortex in the cognitive domain is a distinctly human trait, which, although well documented (cf. Trevarthen 1990a), appears to be poorly understood. For the kind of social cognition and mirroring of movements involved in what I have identified as learning by altercentric participation, an asymmetric and interlinked dual systems architecture appears to be required. There would have to be bi-lateral communication between the system that give rise to body scheme evoked in proprioception and execution of motor programmes and the system that gives rise to the virtual companion scheme, enabling altercentric mirroring.
Preliminary conclusion
Above and elsewhere (essay no 4, this volume) I have attempted to indicate that altercentric participation may be operative in speech perception by the preverbal learner and by the conversational participants in discourse, virtually taking part in one another's production and processing by mirroring the reverse processes in the other (Bråten 1974). The latter is consistent with Liberman's (1957, 1993) finding that speech code perception entails postdiction in the listener of the oral-vocal movements producing the speech. With regard to the preverbal mind I have previously ventured the prediction that an altercentric system, perhaps even altercentric neurons, would be found in humans, when sensitized in the typical and frequent face-to-face interaction with adults. .
It now appears that the above finds independent neurobiological support in current brain research. I have referred to the pertinent discoveries of allocentric cells in rats and of mirror neurons in the primate brain which may be precursory to the kind of system supporting altercentric mirroring in humans. The existence of a mirror system in the human brain, described by Rizzolatti and Arbib (1998), suggests that parts of the neurosociological basis for altercentric mirroring and participation may have been uncovered. Such a mirror system would most likely be a part of the system that subserve altercentric perception in humans, and if sensitized altercentric neurons will be found in humans I would expect them to recruit from a particular human class of mirror neurons.
Altercentricity is probably linked to another human trait enabled by the virtual alter-mechanism complementing ego-processes. This is the dialoguing of voices in the mind, entailing complementary systems that give rise to internal dialogue between crossing perspectives, and which I take to be the unique and defining characteristic of modern human consciousness.
1 Notes for a public lecture 17 October 1997 in connection with a ZiF symposium on Culture in Social Change at University of Bielefeld (opening the research project headed by Günter Dux on 'Theorie des sozialen Wandels' at the Centre for Interdisciplinary Research (ZiF)), drawing upon my commentary in Behavioral and Brain Sciences (1993), vol.16: 515. Some notes about possible neurobiological support have been added to the last part, in preparation for a plenary talk at the conference Mirror Neurons and the Evolution of Brain and Language, Hanse Institute for Advanced Studies, Delmenhorst 5-8 July 2000.
2 Wild chimpanzees living today in West and Central Africa appear to exhibit significant variations, for example, in their use of leaves and various objects, for example in fishing for ants and use of sticks, in their pounding of food, in their reactions to start of rain; some even perform some sort of "dancing in the rain" (Whiten et al. 1999). This variety of patterns, revealed by Whiten, Goodal, Boesch and others, synthesing observations from 151 years of chimpanzee observations suggests cultural transmission, including learning by imitation. Unlike the late Miocene terrestrial apes, venturing out on the nonwooded savanna and coming to evolve a bipedal gait, the chimpanzee (Pan troglodytes) have remained in tree forests (now threatened by the forest extinction).
3 Early infant capacity for imitative learning by virtue of altercentric mirroring may have enabled individuals to modify the learning contexts of intergenerational transmission entailing new selective advantages for future generations. The evolved other-oriented mechanism for learning by altercentric participation may later in phylogeny with the advent of speech have facilitated early learning of articulatory vocal gestures as precursory constituents of speech perception.
4 One may perhaps also speculate about whether there may have been an intervening period in which early hominids lived in river delta landscapes, perhaps even were waders and keeping their offsprings at hand in the water, safe from water-shy land predators. Traces of such settlements may be difficult to find, but if they indeed lived for prolonged period in such conditions, then again this may have entailed the distinct selective advantage of enhanced child capacity for rapid imitative learning in face-to-face interaction..